

1. Fosfoglicéridos
2. Esfingolípidos
3. Esteroles
4. Distribución
5. Asimetría
La organización y propiedades de las membranas celulares está determinada por las características de sus componentes, lípidos, proteínas y carbohidratos. Sin embargo, la diversidad (hay más de mil tipos de lípidos diferentes) y su organización espacial (formando un bicapa) hacen a los lípidos esenciales. Así, ellos definen las propiedades físicas de las membranas. La longitud y el grado de saturación de sus ácidos grasos regulan la fluidez y el grosor de la membrana, y su distribución desigual crea asimetría en las membranas. En la membrana plasmática las cargas asociadas a sus partes hidrofílicas contribuyen a crear un gradiente eléctrico entre la cara externa y la interna, y por tanto a modular el potencial eléctrico de la membrana. Mediante interacciones electroquímicas son capaces de modular la actividad de las proteínas de membrana. Se ha postulado que las interacciones moleculares entre ciertos lípidos producen la segregación de dominios espaciales y funcionales en áreas restringidas de la membrana que afectan también a la localización de las proteínas y a sus funciones. Son las denominadas balsas de lípidos o "lipid rafts". Pero además pueden actuar como segundos mensajeros que abandonan la membrana, viajan a compartimentos intracelulares y desencadenan respuestas celulares.

Los lípidos constituyen aproximadamente el 50 % del peso de las membranas, con unos 5 millones de moléculas por µm2. Las membranas celulares de una célula eucariota contienen más de 1000 tipos de lípidos que aparecen en distinta proporción según el tipo de membrana que estemos considerando. Se estima que aproximadamente el 5 % de los genes de una célula están dedicados a producir sus lípidos.
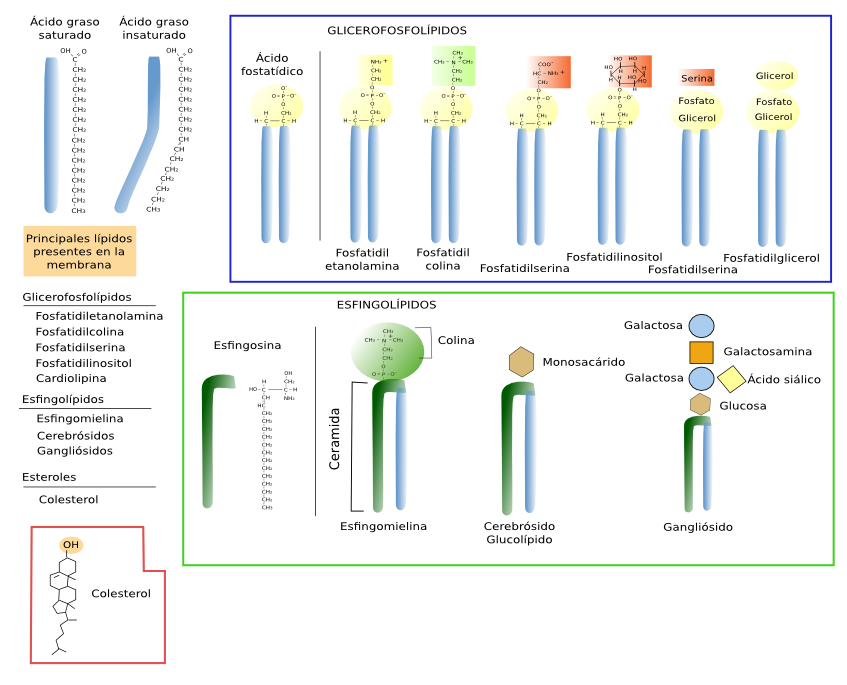
Los lípidos de membrana se caracterizan por poseer una parte apolar o hidrófoba que constituye la parte interna de la membrana y por una parte hidrofílica que está en contacto con el medio acuoso formando las dos superficies de la membrana. Por ello se dice que son moléculas anfipáticas. Se clasifican en tres grupos según su estructura y composición molecular: glicerofosfolípidos (también denominados glicerolípidos, fosfoglicéridos o simplemente fosfolípidos), los esfingolípidos y los esteroles.
1. Fosfoglicéridos o glicerofosfolípidos
Son los lípidos más abundantes ya que representan más del 70 % de los lípidos de las membranas celulares. Estructuralmente constan de tres partes: dos cadenas de ácidos grasos, una molécula de glicerol y un grupo fosfato al que se unen moléculas de diversa naturaleza y que aportan gran parte de la variabilidad de estos lípidos (Figura 1). Las cadenas de ácidos grasos contienen de 13 a 19 átomos de carbono de longitud. La mayoría de los enlaces entre estos carbonos son simples y por tanto se dice que son enlaces saturados. Sin embargo, más de la mitad de estos ácidos grasos tienen al menos un doble enlace entre dos átomos de carbono. Hablamos entonces de ácidos grasos insaturados. Los dobles enlaces hacen que la cadena de ácido graso se doble y, aunque restrinja las posibilidades de movimiento de la cadena, un aumento de la proporción de estos dobles enlaces aumenta la fluidez de la membrana puesto que provoca más separación entre moléculas. Los ácidos grasos constituyen la parte hidrofóbica (fobia por el agua) de los glicerofosfolípidos y son los que constituyen la parte interna de las membranas.

El glicerol hace de puente entre los ácidos grasos y la parte hidrofílica (apetencia por el agua). Esta parte hidrofílica está formada por un grupo fosfato al que se pueden unir una variedad de moléculas, tales como la etonalamina, colina, serina, glicerol, inositol, el inositol 4,5-bifosfato, etcétera. Estos componentes son los que dan nombre a los distintos tipos de glicerofosfolípidos. El tipo fosfatidilcolina representa más del 50 % de los fosfoglicéridos en las membranas eucariotas.
2. Esfingolípidos
Deben su nombre a que poseen una molécula de esfingosina, un alcohol nitrogenado con una cadena carbonada larga, a la cual se le une una cadena de ácido graso, formando la estructura básica denominada ceramida (Figura 2 y ↗ >figura ). A la ceramida se le une una parte hidrofílica que puede ser de diversa naturaleza. Por tanto queda una estructura similar a la de los glicerofosfolípidos, dos cadenas hidrofóbicas unidas a una estructura hidrofílica. Los esfingolípidos constituyen la mayoría de los denominados glicolípidos de las membranas, presentes mayoritariamente en las células animales, es decir, lípidos que poseen uno o más azúcares unidos formando parte de su zona hidrofílica. Otro tipo de esfingolípidos son las esfingomielinas que poseen una etanolamina o una colina fosforiladas en sus zonas hidrofílicas. Los esfingolípidos son más abundantes en las membranas plasmáticas que en las de los orgánulos, y se les propone como lo principales responsables, junto con el colesterol, de la segregación lateral de la membrana en dominios moleculares (balsas de lípidos).

3. Esteroles
El colesterol (Figura 3) es el esterol más importante de las células animales y el tercer tipo de lípido más abundante en la membrana plasmática (hasta el 25 % del total de lípidos), mientras que aparece en pequeñas proporciones en las membranas de los orgánulos celulares como las del retículo endoplasmático (1 %), mitocondrias y lisosomas. El colesterol no aparece en las membranas de las plantas, en algunas células eucariotas, ni en las bacterias, pero estas células tienen otro tipo de esteroles. Los esteroles son esenciales para la integridad y funcionamiento de las membranas eucariotas. Sirven para modular la rigidez, la fluidez y la permeabilidad. Además, contribuyen también a modular la actividad de los receptores acoplados a proteínas G y facilitan la transducción de señales y el tráfico vesicular. El colesterol se localiza entre las cadenas de ácidos grasos de los otros lípidos. Es importante para la organización de la membrana, sobre todo la plasmática, puesto que junto con los esfingolípidos parece contribuir a formar heterogeneidades laterales. También participa en ciertos procesos metabólicos vitales como la síntesis de hormonas esteroideas o de sales biliares, entre otras.

4. La distribución de lípidos depende de la membrana
Aunque hay tres grandes grupos de lípidos de membrana (glicerolípidos, esfingolípidos y esteroles), hay miles de tipos moleculares diferentes de lípidos distribuidos por las membranas celulares (Figuras 4 y 5). La composición de lípidos varía entre las membranas de los diferentes compartimentos membranosos de la célula. Se propone que la propia identidad de los orgánulos viene determinada por la composición de sus membranas, tanto proteínas como lípidos. Por ejemplo, la membrana plasmática tiene una composición lipídica diferente a la membrana del retículo endoplasmático o a la del aparato de Golgi. Estas diferencias se mantienen a pesar del flujo constante de lípidos desde sus compartimentos de síntesis, principalmente el retículo endoplasmático, hasta otras membranas como la plasmática o los endosomas, estando este transporte mediado por vesículas, transportadores y contactos directos entre membranas.


La cantidad y tipo de lípidos varía entre membranas (Figura 5). Por ejemplo, aunque todas la membranas tienen fosfatidil colina, este glicerolípido es más abundante en las membranas del retículo endoplasmático. En la membranas post-Golgi, es decir, membrana plasmática y endosomas, la concentración de esfingolípidos y colesterol es mayor que en el retículo y en las membranas del aparato de Golgi. Las mitocondrias tienen, aparte de otros más extendidos, lípidos de membrana propios como el fosfatidilglicerol y la cardiolipina, que sintetizan ellas mismas.
Esto significa que deben existir mecanismos de segregación por compartimentos membranosos de las diferentes especies de lípidos de membrana. Se han propuesto diversos mecanismos que contribuyen a esta distribución desigual de lípidos:
Síntesis diferencial. La concentración de determinadas especies de lípidos viene condicionada por su lugar de síntesis. Por ejemplo, los esfingolípidos se terminan de ensamblar en el aparato de Golgi y son los compartimentos post-Golgi donde más abundan, pero están ausentes en el retículo endoplasmático. La fosfatidilcolina se sintetiza en el retículo endoplasmático y es ahí donde más abunda. Sin embargo, no siempre es cierto puesto que, por ejemplo, el colesterol se sintetiza en el retículo endoplasmático, pero es más abundante en membranas post-Golgi. Esto es debido a que es transportado rápidamente a otras membranas. Además, durante los procesos de síntesis de unos lípidos se emplean a otros como donantes de partes moleculares por lo que a la vez que se sintetiza una nueva especie lipídica desaparece otra, y todo contribuye a cambiar la composición lipídica de la membrana de ese compartimento.
Selección y transporte. Los lípidos deben ser transportados entre membranas puesto que no difunden libremente por el citosol. Ello implica que el transportador puede seleccionar qué lípidos acarrea de un lado a otro cambiando así las proporciones de determinadas especies de lípidos tanto del compartimento fuente como del de destino. Estos transportadores son fundamentalmente las vesículas, las cuales forman sus membranas con lípidos de las propias membranas del compartimento de partida. Por ejemplo, las vesículas que van desde el Golgi a la membrana y a los endosomas están enriquecidas en esfingolípidos y colesterol, respecto la concentración de estas moléculas en las propias membranas del Golgi. También hay transportadores de lípidos individuales que son proteínas que son capaces de extraer un lípido de una membrana, transportarlo por el citosol y colocarlo en otra membrana.
Contactos entre membranas. Las membranas de determinados orgánulos pueden observarse extremadamente próximas. Hay proteínas que se colocan entre las membranas de dos orgánulos cuando están muy próximas espacialmente, haciendo de puente para el trasiego de lípidos entre las membranas de ambos orgánulos. Esto ocurre entre membranas de compartimentos que no están comunicados mediante vesículas, por ejemplo, entre el retículo endoplasmático y las mitocondrias. Pero también entre compartimentos comunicados por vesículas como el retículo endoplasmático y el lado trans del aparato de Golgi, entre el retículo endoplasmático y los endosomas, o entre el retículo endoplasmático y la membrana plasmática. Por ejemplo, el complejo proteico CERT (ceramide transfer protein) transfiere ceramida desde las membranas del retículo endoplasmático al lado trans del aparato de Golgi.
Degradación y reciclado diferencial. Todas las membranas sufren un proceso de reciclado de lípidos, bien por degradación in situ de los lípidos o por su salida en vesículas de reciclado. En ambos procesos es posible un mecanismo de selección de unos tipos de lípidos respecto a otros que condicionará la población de lípidos de la membrana.
5. Asimetría
La asimetría es la distribución diferencial de lípidos entre las dos hemicapas que componen las membranas. Mientras en el retículo endoplasmático ambas hemicapas son muy similares, pero no idénticas en cuanto a la composición lipídica, las membranas del lado trans del aparato de Golgi y los compartimentos post-Golgi suelen tener una clara diferencia de composición entre monocopas. Para saber más: ↗.
-
Bibliografía ↷
-
Bissig C, Gruenberg J. 2013. Lipid sorting and multivesicular endosome biogenesis. Cold Spring Harbour perspectives in biology. 5:a016816.

Janmey PA, Kinnunen PKJ. 2006. Byophisical properties of lipids and dynamic membranes. Trends in cell biology. 16:538-546.
Vance JE. 2015. Phospholipid synthesis and transport in mammalian cells. Traffic. 16:1.
van Meer G, Voelker DR, Feigenson GW. 2008. Membrane lipids: where are they a how they behave. Nature reviews in molecular cell biology. 9:112-124.
-
 Membrana celular
Membrana celular 
{kind=link}