

1. Introducction
In the plant embryo, nearly all cells can divide and subsequently differentiate into functional mature cells. However, as the plant grows, clusters of undifferentiated cells remain in some regions of the body of the plant. In addition, groups of differentiated cells may become undifferentiated. Meristems are populations of undifferentiated cells present in the plant body with the ability to proliferate and differentiate into a variety of functional cells. Meristems are responsible for the growth of the plant, and they may be present during the plant's lifespan. Not all proliferating cells in the meristem end up as differentiated cells. Within the meristem, some cells known as initial cells remain undifferentiated after mitosis, leading to a permanent pool of undifferentiated cells that preserve the meristematic features as long as the plant is alive. The inner environment of the meristem has signaling molecules that promote proliferation and maintain undifferentiation.
Meristems may be permanent and functional throughout the plant's lifespan, as happens to the shoot and root apical meristems. Others are transient and active for a short period of time, such as those giving rise to the leaves, flowers, and petioles. Some meristems may appear quite late after germination, such as the phelloderm and vascular cambium.
Meristematic cells show the cytological features of undifferentiated cells: they are small and isodiametric, and show a very thin primary cell wall. The cytoplasm is rich in ribosomes, proplastids, and many small vacuoles, but but has a low content of endoplasmic reticulum and inclusions. A well-developed Golgi complex synthesizes the components of the cell wall. A large nucleus contains much condensed chromatin and is found centrally in the cell. Meristematic cells divide by mitosis and may be totipotent, meaning they are able to differentiate into the entire variety of cell types of a mature plant, or pluripotent when giving rise to some cell types. Plants grow by producing new cells and by the growth of these new cells, which is accomplished by the accumulation of water in their vacuoles. Normally, meristematic cells are densely packed with almost no intercellular spaces.
Almost all meristems contain a region called the niche, which contains the initial cells (stem cells), surrounded by progenitor cells. Progenitor cells divide many times and differentiate into functional cells. The final cell type into which a meristematic cell will differentiate depends on its spatial position within the meristem and the molecular signals available in that place. Although the differentiation of meristematic cells is influenced by the cell lineage they belong to, a particular location inside the meristem is a stronger signal that finally determines the differentiation pathway. The initial cells give rise to progenitor cells, which first form a proliferation zone with a high division rate. Then cells stop dividing and increase their size, generating the elongation zone. Finally, cells differentiate into a particular cell type of the organ.
The classification of meristems is based on their topographic position in the plant's body and on the time they become active along the plant's lifespan (Figure 1).

2. Primary meristems


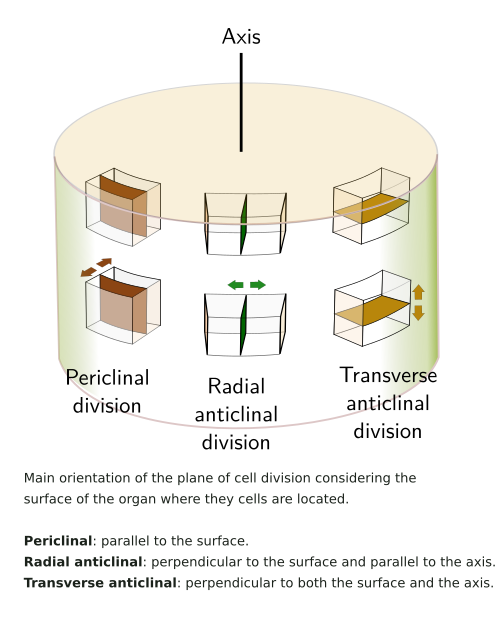
Primary meristems are first observed in the embryo, and their activity results in so-called primary growth, which is mainly involved in lengthening the plant. During this activity, cells undergo anticlinal cell divisions, meaning that the plane of the cell division is perpendicular to the surface of the plant body (see figure). Primary meristems are able to form new organs. Primary meristems are found in the tips of shoots (Figure 2) and main roots, and hence they are called shoot (or caulinary) apical meristem and root apical meristem, respectively. All cells of the shoot derive from the shoot apical meristem during primary growth, including leaves, flowers, and branches. The axillary buds, from which stem branches arise, are also derived from the shoot apical meristem. However, the lateral roots are generated from the endodermis, a tissue also derived from the root apical meristem. An apical meristem is present in every shoot branch and lateral root. The shoot apical meristem is covered and protected by the youngest leaf primordia, whereas the root apical meristem is wrapped by the calyptra.
The number of cells constituting an apical meristem is relatively constant throughout the life of the plant, since there is a balance between the cells that leave the meristem and form tissues and the new cells that are generated within the meristem. In the shoot apical meristem, there is a central zone and a central organizer, which form the niche that sustains the initial cells, whereas in the root apical meristem, the niche is the quiescent center, with initial cells surrounding it. Three lineages of cells are formed in the apical meristems. The outermost layer is the protoderm, which differentiates into the epidermis; then comes the procambium, which gives rise to the primary vascular tissues (primary xylem and primary phloem); and finally the ground meristem, the precursor of ground tissues (parenchyma, collenchyma, and sclerenchyma).

Plant cells do not move, and this makes it possible to predict what the fate will be of a cell by knowing the spatial location within the meristem. In the apical meristems, while the central zone produces new cells, the meristem moves away, and these new cells fall under the influence of molecules that guide their differentiation process. This separation between the meristem and new cells is actually a consequence of the continuous mitotic activity of the meristematic cells, which push the meristem away.
The axillary buds are not associated with the leaves, despite it appears otherwise. In fact, axillary buds develop in the axil of the second or third leaf from the apex. It means that they appear later than the leaves. Axillary buds arise as small cellular populations that detach from the apical meristem. However, their final differentiation is induced by the leaf, as the removal of the leaf prevent axillary bud formation.
3. Secondary meristems


Secondary, or lateral, meristems are present in those plants that grow in thickness, i.e., they undergo secondary growth (Figure 1). These meristems arise later on during the growth of the body of the plant and are responsible for producing wood and bark. They increase the diameter of shoots and roots, and their cells divide by periclinal division (see figure). Secondary meristems are typical of gymnosperms and dicots. They are not found in most pteridophytes and monocots, nor in some organs such as leaves. Plants with secondary growth also perform primary growth at the ends of shoots and roots. There are two types of lateral meristems: the vascular cambium, which differentiates from the procambium and produces secondary vascular tissues (secondary xylem and secondary phloem), and the cork cambium (phellogen), which arises from various parenchymatous tissues in the cortex and produces phelloderm inward and cork outward. Both meristems are arranged along shoots and roots forming a tube-like structure, sometimes with gaps in the wall. The vascular cambium is found between the secondary xylem and the secondary phloem, whereas the cork cambium is located between the phelloderm and cork. The vascular cambium is formed from the procambium and from new meristematic cells differentiated from the interfascicular parenchyma. In the roots, the vascular cambium arises from the procambium and the pericycle.
4. Other meristems
In monocots, the elongation of shoots primarily relies on intercalary meristems rather than the apical meristems Intercalary meristems are meristematic tissues originating from the apical meristem. They continue the proliferative activity at some distance from the place where they originated, i.e., they are inserted (intercalated) among tissues that are no longer meristematic. The best-known examples of intercalary meristems are those located in the stem internodes (mainly at the base of the internodal region) and in leaves, particularly in grasses. Close to the shoot apical meristem, the nodes are very close to one another. During development, the nodes are separated from each other by cell proliferation and elongation. In dicots, the separation of the nodes is mostly done by cell elongation.
There are two additional meristems play a rol in the development of the vascular system: provascular and preprocambium. The provascular tissue is an embryonic tissue located in the prospective vascular cylinder. Although this provascular meristem does not directly give xylem or phloem, it produces precursors of the procambium, which give rise to xylem and phloem. Preprocambium cells are found in leaves and are the precursors of procambium. They cannot be distinguished from the ground tissue cells. In the leaves, the procambial cells are recruited from a subepidermal population of the leaf primordia. These cells also differentiate into parenchyma cells. The meristematic cells of the leaf are formed under the influence of a high concentration of auxin. They arrange into strands that become the leaf veins, which grow by the addition of new cells instead of division and elongation. The mechanisms of auxin canalization lead to this behavior because auxin is conducted toward neighboring cells, which dedifferentiate into procambial cells.
Meristems may arise by dedifferentiation of parenchyma, collenchyma, and pericycle (in roots). For instance, it was already mentioned that a part of the vascular cambium is formed from the interfascicular parenchyma in the stem. Other instances of dedifferentiation include epidermal cells coming from parenchyma, or phloem, or the adventitious roots that emerge after dedifferentiation of stem functional tissues.
In monocots, the secondary growth was lost during evolution. This might have occurred due to all cells of the procambium differentiating into vascular tissues, leaving no vascular meristem remaining. In these plants, the growth in thickness is due to an increase in cell size. However, palms and other tree-like monocots (some asparagales) show very thick stems by enlarging the parenchyma tissue and increasing the number of vascular bundles in regions quite distant from the stem tip. This type of growth is known as anomalous growth, and it is produced by the activity of the peripheral growing meristem. In palms, this meristem, referred to as the secondary growing meristem, is found more superficially than the vascular bundles. It gives parenchyma outward, and vascular bundles and parenchyma cells inward. In some monocots, this peripheral meristem may extend to the roots as a cylindrical structure, resembling a vascular cambium. The activity of this meristem produces tissue similar to the secondary tissues when that part of the stem stops growing in length. Curiously, it produces most of the tissue toward the inner region of the stem.
-
Bibliography ↷
-
Augstein F, Carlsbecker A. 2018. Getting to the Roots: A Developmental Genetic View of Root Anatomy and Function From Arabidopsis to Lycophytes. Frontiers in plant sciences. 9: 1410. DOI: 10.3389/fpls.2018.01410.
Miyashima S, Sebastian J, Lee J-Y, Helariutta Y. 2013.Stem cell function during plant vascular development The EMBO journal. 32: 178-193. DOI: 10.1038/emboj.2012.301 .
Truskina J, Vernoux T. 2018. The growth of a stable stationary structure: coordinating cell behavior and patterning at the shoot apical meristem. Current opinion in plant biology. 41: 83-88. DOI: 10.1016/j.pbi.2017.09.011.
-
 Introduction
Introduction 
{kind=link}