

In plants, it is challenging to distinguish between excretion and secretion processes since the byproducts of both are usually stored within the same plant compartments. In general, secretion is the accumulation of secondary products resulting from metabolism that are not immediately utilized, along with primary metabolites that are going to immediately participate in metabolic reactions in the cell. Secretory cells arise from the differentiation of epidermal or parenchyma cells and do not form true tissues. The structures responsible for secretion exhibit highly diverse morphologies and are located in a variety of regions of the plant. In fact, they can be found in both internal and superficial parts of the plant and can be either a single secretory cell or a multicellular secretory structure. In addition, they can produce a wide variety of products.
The waste products are accumulated in particular cells, transferred to some tissues, sometimes to dead tissues, as well as into cavities and ducts of the plant. For instance, many waste products are stored in the wood.
Here, we are going to classify secretory structures according to their location in the body of the plant: superficial or internal.
1. Superficial secretory structures
There are many secretory cells in the peripheral tissues of plants, either as unicellular and multicellular glandular trichomes in the epidermis or as components of the epidermal layer. These secretory cells arise by division and differentiation of epidermal cells. Some of them mainly release hydrophilic or lipophilic substances, while others release both hydrophilic and lipophilic compounds. Superficial secretory cells are found in stems, leaves, fruits, and flowers. For example, there is abundant secretion in petals, where the released molecules cause the fragrance of flowers. Although these cells are highly specialized in secretion, all epidermal cells are actually secretory, since all of them release substances that are deposited on their walls or released into the environment.
Hydathodes are structures that release water along with some substances from the interior of the leaf to its surface. This process is called guttation and occurs due to water pressure coming from the roots.
Hydathodes are modifications of some parts of the leaves and are located along the margins or at the tip of the leaf (Figures 1 and 2). Structurally, hydathodes consist of a) terminal tracheids of leaf nerves, b) the epithem, which consists of parenchyma cells showing thin-walls and few chloroplast, and located at the leaf nerve endings, c) a sheath or envelope that is continuous with the epidermis (the sheath cells may be suberized and may even have Caspary bands), and d) a watery opening or pore (watery pores are small nonfunctional stomata that have lost the ability to regulate opening and closing). Although this is the general organization, there may be variations, such as the lack of the sheath or the epithem, or it might also feature an opening that is not a stoma.


Although hydathodes typically occur at the margins and tips of leaves, they also appear on the surface in some species, referred to as laminar hydathodes. Hydathodes are generally associated with the release of water from the plant tissues, yet in many xerophytic species they are specialized for the absorption of condensed fog or dew water, i.e., the reverse process. Some authors have described the so-called active hydathodes, also known as trichome-hydathodes, as glandular trichomes that release water. These structures would release water without being influenced by the osmotic pressure. Hence, they are considered active incontrast to those that would be passive.
Nectaries are secretory structures that release sugar-rich solutions made from the substances supplied by the phloem. Two categories can be distinguished: floral nectaries, which are directly associated with pollination, and extrafloral nectaries, which are found on the vegetative parts of the plant. Nectary structures vary greatly, from simple epidermal glandular surfaces to more complex structures.
Osmophores are secretory structures that produce the fragrance of flowers by releasing aromatic volatile substances.
Salt glands are secretory structures found in halophyte plants (those living in high salinity environments). These glands store and release ions to regulate the salt levels within the plant. The salt cells are modified multicellular trichomes (Figure 3). About 0.25% of flowering plants are regarded as halophytes. Salt glands increase the tolerance to salinity. However, the cellular and molecular mechanisms may differ as salt glands were developed independently by different plant taxa. They are an example of convergent evolution. The salt glands are found in the epidermis, mostly in leaves, of plants that thrive in dry soil, swamps, and mangroves with high salinity. Salt ions move from the xylem to the parenchyma by apoplastic pathways and reach the cells that support the salt gland in the epidermis. From these basal cells, salt travels via symplast crossing plasmodesmata and gets to the salt storage cells, where it is transferred to the vacuole. As these cells disintegrate, salt is released as white deposits over the epidermis of the plant. The salt precipitates do not enter plant tissues again due to the epidermal cuticle. In tamarinds, the salt is transferred to vesicles of the secretory cells that subsequently merge with the plasma membrane. From the pericellular space, salt gets to the epidermal surface through small channels present in the cuticle.

There are two types of salt glands regarding their functionality: those releasing outside of the plant and those that store ions in specialized vacuoles. According to their cellular organization, four types of salt glands have been described (Figure 3). Type I: salt vesicle, in which a large cell stores the salt. Type II: multicellular with 4 to 40 cells consisting of collector and secretory cells covered by a cuticle. Type III: bicellular filliform structures with a basal and an apical cell. Type IV: vacuolar unicellular or filliform secretory. Types I and II are found in dicots, while types III and IV are present in monocots. The type I is a very large cell with a vacuole, which can subsequently be broken to release its content. This cell may be found over a group of epidermal cells. Type II is multicellular with two cell populations: basal collector and superficial secretory. Secretory cells are covered by a cuticle, which may be perforated. Salt is accumulated between the plasma membrane and the cuticle. These glands show bulb-like or disc-like shapes, usually forming a depression in the epidermal layer. Salt is usually released into the center of these structures. In some glands, a third type of cell is found connecting the collector and releasing cells. Type III glands share features with the secretory glands of dicots but are structurally simpler. They are made up of one or two cells, with or without a cuticle, and with characteristic basal cells. Type IV glands are present in the wild rice and are unicellular trichomes.
Glandular trichomes are external secretory structures present in about 30% of plant species (Figure 4). For instance, the fragrance of plants mainly arises from the volatile substances released by these structures. Glandular trichomes are commonly multicellular, with glandular cells at the distal end attached to the epidermis by non-glandular cells known as basal cells and by other cells forming a stalk or peduncle. However, some trichomes are unicellular. Glandular cells have a primary cell wall, sometimes covered by a cuticle at the joining area with the stalk cells, and contain a cytoplasm with abundant organelles such as mitochondria, endoplasmic reticulum, and Golgi cistern stacks. However, there are differences between glandular cells among plant species. For example, glandular cells releasing terpenes contain very scarce Golgi apparatus, or it is not visible at all.

Glandular trichomes contain cells that release substances to the environment with different functions, either as volatile substances or as those that remain on the surface of the plant. Glandular cells can synthesize the molecules to be released, even though they sometimes have the ability to perform photosynthesis too. It means that some glandular cells may live as rather independent units. However, in other cases, the molecular building blocks for synthesizing the compounds to be released are coming from the underlying plant tissues through the stalk cells of the trichome.
Glandular trichomes are classified according to the compounds they release or their morphological features. There are trichomes with a basal cell, one or more in the stalk, and a few secretory cells at the distal end. These trichomes commonly release relatively non-volatile substances that are laid on the surface of the plant. Other types of trichomes show a basal cell, a short cell in the stalk, and a head with one or more secretory cells, showing a large cavity between the cuticle and the primary cell wall, which is filled with substances. There are many other examples of glandular trichomes. Different types of trichomes can be found in the same plant, even in the same organ. For example, in carnivorous plants, there are trichomes for fetching the insect by releasing mucilaginous substances or nectar, and others release proteolytic enzymes for digestion. The number of trichomes, both glandular and not glandular, is influenced by the environment. For instance, it may increase after the attack of a herbivore.
The release of secretory compounds may be achieved in different ways. Those secretory products stored between the cell wall and the cuticle are released when the trichome is touched and the cuticle is detached. In other cases, the cuticle has tiny channels to guide substances to the environment. Some glandular cells store the substances in their cytoplasm so that they are released when the cell breaks. Initially, the precursor molecules arrive into secretory cells via the stalk cells of the trichome, but the final secretory products synthesized in the secretory cells do not travel back to the plant organ. This is because the mature secretory cells get a thicker cuticle, and the cells of the stalk close to the secretory cells work now as a barrier.
There is a wide variety of glandular trichomes regarding the compounds they release. Some synthesize and release organic acids, others release salt, mucilage (polysaccharides), terpenes, combinations of terpenes and mucilage, nectar, proteolytic enzymes by trichomes of carnivorous plants, among others. Some of these substances may serve as antimicrobial compounds. One important function of glandular trichomes is mediating interactions between plants and animals. For instance, released terpenes may attract insects or be toxic to them, produce alarm, regulate insect development, or have other effects depending on the insect species. More curiously, some terpenes attract insect predators or insect parasites. This last strategy is known as indirect defense.
What are sources of energy and carbon for the production of secretory products and metabolites in glandular trichomes? They are mostly found within the plant body. Even in those glandular trichomes that are able to perform photosynthesis, the energy of the photosynthetic process is not employed for fixing CO2, but for metabolic pathways involved in secretion. The carbon sources of glandular trichomes are saccharides synthesized in other cells of the body plant, but the characteristic metabolites of glandular cells are synthesized by the glandular trichome itself.
2. Internal secretory structures
Internal secretions are products stored within plant tissues, sometimes throughout the lifespan of the plant. Internal secretory structures are far from the epidermis and primarily found in the cortical parenchyma of stems, leaves, roots, and fruits. We can distinguish three types of internal secretory structures:
Secretory cells. They are isolated cells that differ from the surrounding cells and may be from isometric to enlarged forms resembling sacs or tubes. These are specialized cells referred to as idioblasts. Many secretory cells contain mixtures of substances, yet the contents remain unidentified in many other secretory cells. They synthesize and store a wide variety of products, such as resins, mucilage, tannins, oils, gums, and even crystallized substances. Crystallized precipitates are often observed in the cytoplasm, which correspond to calcium oxalate. These crystals are found in cells of the parenchyma, secondary phloem, and in some cells of the stratified epidermis (litocysts). They are named according to the crystal morphology: druses are spherical or star-like (Figure 5), raphides resemble needles, styloids are acicular in shape, and prismatic crystals are also found. The function of calcium oxalate crystal in the plant cells is no yet fully understood.

Secretory cavities and ducts (canals) differ from secretory cells because they release substances into intercellular spaces. Secretory cavities are small secretory spaces, whereas secretory ducts are large secretory spaces. These spaces may be found anywhere in the plant and can be formed by two different cellular processes: schizogeny and lysogeny. They are commonly formed by the separation of cells of vascular tissues or ground tissues, resulting in an intercellular space lined with secretory epithelial cells. These spaces are called schizogenous secretory cavities and ducts, as is the case with most resin ducts. They can also be originated by the disintegration and dissolution (autolysis) of the glandular cells. In this case, the secretory product is formed in the cells that eventually break down, but the compounds remain in the resulting space. These spaces are known as lysogenic spaces, like the lysogenic cavities in citrus fruits. Some authors also recognize a third pattern of development, the schizolysogeny. The formation of schizolysogenous cavities and ducts is initially schizogenous (separation of intact cells) but followed by lysogeny at later stages, when the epithelial cells lining the space undergo autolysis, further enlarging the space. The resin ducts in the bud scales of Pinus pinaster are an example of ducts following a schizolysogenous pattern of development (see figure).

Gymnosperms contain internal secretory structures that release a compound known as resin. These structures may be arranged as resiniferous cavities, as seen in spruces and cedars, or as resiniferous ducts, like in pines. When the resin is exposed to the air, it is transformed into a protective physical and chemical barrier. Resiniferous cavities and ducts are formed by secretory cells with very thin cell walls.
Laticifers refer to individual cells or groups of connected cells containing a liquid called latex. When formed of several cells, they can produce complex tube-like structures. According to the structure, they can be classified into articulated and nonarticulated laticifers. Articulated laticifers are made up of longitudinal chains of connected cells, whereas the nonarticulated laticifers consist of long tube-like single cells. Non-articulated laticifers are developed in the embryo stage, where precursor cells are already present. Articulated laticifers are formed from the apical meristems. However, both end up with a similar set of ducts showing a branching pattern that varies according to the plant species. Laticifers are found in a wide variety of species, ranging from herbaceous to woody plants. Laticeferous cells do not show secondary cell walls. The term "latex" refers to a heterogeneous secretory substance that greatly varies in appearance (color) and composition (it may contain substances like carbohydrates, organic acids, polyterpenes, alkaloids, salts, sterols, fats, mucilage, proteins, and starch), thus resulting in many types of latex.
-
Bibliography ↷
-
Dassanayake M, Larkin JK. 2017. Making plants break a sweat: the structure, function, and evolution of plant salt glands. Frontiers in plant science. 8: 406. DOI: 10.3389/fpls.2017.00406,
Lange BM. 2015. The evolution of plant secretory structures and emergence of terpenoid chemical diversity. Annual review of plant biology. 66: 139-159. DOI: 10.1146/annurev-arplant-043014-114639.
-
 Protection
Protection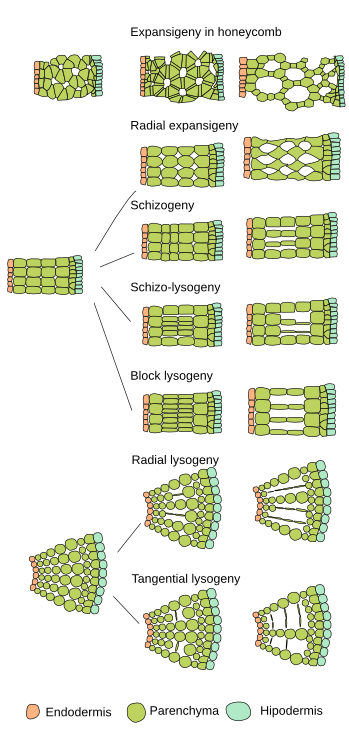{kind=link}